Los métodos de investigación filogenética están orgánicamente relacionados con los métodos de estudio de los hechos de la evolución. Hasta ahora, el método morfológico debe considerarse el principal método de investigación filogenética, ya que las transformaciones en la forma de un organismo siguen siendo el hecho más obvio y nos permiten rastrear los fenómenos de transformación de las especies con un gran grado de éxito.
Por supuesto, de esto no se sigue que otros métodos no sean aplicables a los estudios filogenéticos: fisiológicos, ecológicos, genéticos, etc. La forma y función de un organismo están indisolublemente ligadas. Cualquier organismo se forma bajo la influencia de factores ambientales específicos, interactúa con él y tiene ciertas conexiones con otros organismos. Sin embargo, la forma de un organismo, su estructura, siempre sigue siendo un indicador sensible de todas estas conexiones y sirve como hilo conductor para el investigador de cuestiones filogenéticas. El método de investigación morfológica ocupa una posición de liderazgo en el estudio de la filogenia y sus conclusiones generalmente se han confirmado cuando se prueban con otros métodos. La gran ventaja del método morfológico es la posibilidad de combinarlo con un método comparativo de investigación, sin el cual es imposible detectar el hecho mismo de la transformación de los sistemas vivos. La validez del método morfológico se ve reforzada en gran medida por el hecho de que es esencialmente profundamente autocrítico, ya que puede aplicarse en varias direcciones.
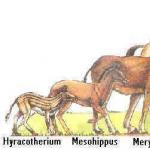
Si disponemos de gran material paleontológico (por ejemplo, la evolución del caballo), podemos aplicar el método morfológico comparativo a series sucesivas de antepasados y descendientes y así identificar las direcciones y métodos de evolución de un grupo determinado. La figura da una idea de la esencia del método morfológico comparativo aplicado a los antepasados del caballo. La reducción secuencial de los dedos laterales y el desarrollo del dedo medio (III) muestran la dirección del desarrollo evolutivo de la “fila del caballo”.
Comparación de las extremidades de los jerbos con un número decreciente de dedos y una especialización creciente. 1 - pequeño jerbo Allactaga elator, 2 - Salpingotas Koslovi, 3 - Dipus sagitta de patas tupidas. I-V - dedos del primero al quinto (según Vinogradov)
Además, los datos paleontológicos están en armonía con los estudios anatómicos comparativos de las formas modernas. La figura compara las extremidades de tres formas con un número de dedos decreciente. Aunque no se trata de una serie filogenética, sí da la impresión de que los tres miembros son el resultado de la manifestación de procesos similares que han alcanzado diferentes etapas de desarrollo. Por tanto, el método morfológico comparativo, en relación con las formas modernas, independientemente de la paleontología, hace probable que, por ejemplo, un pie de un solo dedo se haya desarrollado a partir de uno de varios dedos. Cuando a estas conclusiones se suman los hechos de la embriología comparada, que muestran que, por ejemplo, en el embrión de un caballo se forman los dedos laterales de los pies y luego se reducen gradualmente, entonces nuestra conclusión sobre el origen del caballo de un dedo a partir de muchos El ancestro con dedos se vuelve aún más probable.
La coincidencia de estos datos muestra que los hechos de la paleontología, la anatomía comparada de las formas adultas y la embriología comparada se controlan y complementan mutuamente, formando en su totalidad un triple método sintético de investigación filogenética propuesto por Haeckel (1899) y que no ha perdido su vigencia. importancia en la actualidad. Se acepta que la coincidencia de datos de paleontología, anatomía comparada y embriología, en cierta medida, sirve como prueba de la exactitud de las construcciones filogenéticas.
Estos son los principios más generales de la investigación filogenética.
Consideremos ahora los elementos brevemente descritos de un método unificado de investigación filogenética.
Los datos paleontológicos son los más convincentes. Sin embargo, tienen un defecto importante: el paleontólogo se ocupa únicamente de las características morfológicas y, además, de las incompletas. El organismo en su conjunto está fuera del alcance de la investigación paleontológica. En vista de esto, es especialmente importante que un paleontólogo tenga en cuenta todos los signos disponibles de los animales cuyos restos está tratando. De lo contrario, sus conclusiones filogenéticas pueden ser erróneas.
Supongamos que las formas A, B, C, D, E, E se reemplazan entre sí en horizontes geológicos sucesivos, y que el paleontólogo tiene la oportunidad de observar una cierta suma de sus características: a, b, c, etc. Supongamos además que la forma A tiene las características a 1, b 1, c 1, y en las formas B, C, D, ... estas características cambian (respectivamente a 2, b 2, c 2 .. a 3, b 3, c 3..., etc.). Luego, con el tiempo, obtenemos la siguiente serie de datos.
Esta tablilla corresponde, por ejemplo, a una “fila” de ancestros equinos, donde desde Eopippus hasta el caballo tenemos una sucesión en el desarrollo de una serie de características. La tabla muestra el desarrollo sucesivo de todas las características principales. Cada signo posterior (por ejemplo, un 4) se deriva de cada uno anterior (por ejemplo, un 3). En tales casos resulta probable que la serie A, B, C, D, E, E forme serie filogenética, es decir, un número de antepasados y sus descendientes. Esta es la serie desde eohippus hasta horse y algunas otras.
Ahora supongamos que estamos tratando con los siguientes datos,
es decir, enunciamos una serie de formas que se reemplazan sucesivamente en el tiempo, y según uno de los signos (b) obtenemos una imagen del desarrollo secuencial de b 1 a b 5. Sin embargo, nuestra serie no es una serie filogenética, ya que, por ejemplo, respecto a los caracteres ayb no observamos una especialización consistente. Por ejemplo, la especie A tiene la fórmula A (a 1, b 1, b 1), pero la especie B claramente no es su descendiente directa, ya que tiene la fórmula B (a 4, b 2, b 2), etc. Obviamente, aquí Se trata de sucesivos “fragmentos” de un árbol filogenético, del que muchas ramas no han sido encontradas. Por lo tanto, la serie A, B, C, D, D, E es en realidad igual a A, B 1, C 2, D 3, D 1. Esta serie se llama escalonada. Para explicar las diferencias entre ésta y la serie filogenética, utilizaremos una imagen que muestra la evolución del caballo. Aquí serán filogenéticas las siguientes series: eohippus, orohippus, mesohippus, parahippus, merigippus, pliohippus, plesippus, caballo. Por ejemplo, se escalonarían las siguientes series de formas: hyracotherium, epihippus, myohippus, anchytherium, hipparion, hippidium, horse. Todos estos no son ancestros y descendientes, sino ramas laterales sucesivas pero dispersas del árbol filogenético.
Como puede ver, la serie escalonada es de gran importancia laboral, ya que, en base a ella, se podría concluir que el caballo desciende de un ancestro polidáctilo.
Finalmente, puedes encontrarte rango adaptativo, mostrando el desarrollo de cualquier adaptación. Una serie de este tipo puede ser parte de una serie filogenética, por ejemplo, la adaptación de la pata de un caballo a correr, pero a menudo no es así, y se puede crear una serie adaptativa incluso a expensas de formas modernas que no forman una serie filogenética en absoluto. Como vemos, un paleontólogo tiene que afrontar grandes dificultades. Su material es fragmentario e incompleto.
Sin embargo, cierta compensación por lo incompleto de los datos paleontológicos es la posibilidad de extender los datos ambientales a la paleontología. Una determinada forma de un órgano (la estructura de una pierna, la estructura del aparato dental, etc.) permite sacar conclusiones sobre el estilo de vida e incluso la composición de los alimentos de los animales extintos. Esto da lugar a la posibilidad de reconstruir sus relaciones ecológicas. El campo de conocimiento correspondiente, establecido en los trabajos de V. O. Kovalevsky, se llamó paleobiología (Abel, 1912). Compensa la fragmentación de las ideas de los paleontólogos sobre los animales extintos. Respecto a las formas sin esqueleto, la paleontología sólo proporciona material insignificante sobre filogenética, y en estos casos la morfología comparada con su método de estudio comparativo de estructuras homólogas de formas adultas y embrionarias de la modernidad geológica ocupa el primer lugar. La falta de datos paleontológicos hace que sea mucho más difícil sacar inferencias filogenéticas. Por tanto, nuestras construcciones filogenéticas son más fiables en relación con aquellas formas por las que se conoce el material paleontológico.
Sin embargo, incluso en ausencia de datos paleontológicos, el investigador no permanece desarmado. En este caso utiliza otro método, a saber, el estudio de las etapas del desarrollo ontogenético.
Si encuentra un error, resalte un fragmento de texto y haga clic en Ctrl+Entrar.
Pregunta 1. ¿Cuál es la diferencia entre macro y microevolución?
Por microevolución nos referimos a la formación de nuevas especies.
El concepto de macroevolución denota el origen de taxones supraespecíficos (género, orden, clan, tipo).
Sin embargo, no existen diferencias fundamentales entre los procesos de formación de nuevas especies y los procesos de formación de grupos taxonómicos superiores. El término "microevolución" en el sentido moderno fue introducido por N.V. Timofeev-Resovsky en 1938.
Pregunta 2. ¿Qué procesos son las fuerzas impulsoras de la macroevolución? Dé ejemplos de cambios macroevolutivos.
En la macroevolución operan los mismos procesos que durante la especiación: la formación de cambios fenotípicos, la lucha por la existencia, la selección natural, la extinción de las formas menos adaptadas.
El resultado de los procesos macroevolutivos son cambios significativos en la estructura externa y la fisiología de los organismos, como, por ejemplo, la formación de un sistema circulatorio cerrado en los animales o la aparición de estomas y células epiteliales en las plantas. Las adquisiciones evolutivas fundamentales de este tipo incluyen la formación de inflorescencias o la transformación de las extremidades anteriores de los reptiles en alas y muchas otras.
Pregunta 3. ¿Qué hechos subyacen al estudio y la evidencia de la macroevolución?
La evidencia más convincente de procesos macroevolutivos proviene de datos paleontológicos. La paleontología estudia los restos fósiles de organismos extintos y establece sus similitudes y diferencias con los organismos modernos. A partir de los restos, los paleontólogos reconstruyen la apariencia de organismos extintos y aprenden sobre la flora y la fauna del pasado. Lamentablemente, el estudio de las formas fósiles nos ofrece una imagen incompleta de la evolución de la flora y la fauna. La mayoría de los restos consisten en partes sólidas de organismos: huesos, caparazones y tejidos de soporte externos de plantas. De gran interés son los fósiles que conservan rastros de madrigueras y pasajes de animales antiguos, huellas de extremidades u organismos enteros dejados en sedimentos que alguna vez fueron blandos.
Pregunta 4. ¿Cuál es la importancia del estudio de las series filogenéticas?Material del sitio
El estudio de series filogenéticas construidas sobre la base de datos de paleontología, anatomía comparada y embriología es importante para el desarrollo posterior de la teoría general de la evolución, la construcción de un sistema natural de organismos y la reconstrucción de la imagen de la evolución de un grupo sistemático específico de organismos.
Actualmente, para construir series filogenéticas, los científicos utilizan cada vez más datos de ciencias como la genética, la bioquímica, la biología molecular, la biogeografía, la etología, etc.
1. ¿Qué hechos pueden indicar una conexión entre plantas y animales extintos y modernos?
Respuesta. Según la teoría sintética de la evolución, el proceso evolutivo que ocurre en la naturaleza se divide en dos etapas: microevolución y macroevolución.
La macroevolución implica procesos que conducen al surgimiento de unidades sistemáticas más grandes que una especie. Al estudiar la macroevolución, las ciencias naturales modernas han acumulado una serie de hechos científicos que prueban la evolución del mundo orgánico. Cualquier hecho científico que pruebe al menos uno de los siguientes puntos puede considerarse prueba de evolución.
Unidad del origen de la vida (la presencia de características comunes en todos los organismos vivos).
Relaciones entre organismos modernos y extintos o entre organismos de un grupo sistemático grande (la presencia de características comunes en organismos modernos y extintos o en todos los organismos de un grupo sistemático).
La acción de las fuerzas impulsoras de la evolución (hechos que confirman la acción de la selección natural).
La evidencia de la evolución obtenida y acumulada en el marco de una ciencia particular constituye un grupo de evidencia y recibe el nombre de esta ciencia.
La paleontología es la ciencia de los restos fósiles de organismos extintos. Se considera que el fundador de la paleontología evolutiva es el científico ruso V. O. Kovalevsky. La evidencia de la evolución incluye formas de transición fósiles y series filogenéticas de especies modernas.
Las formas de transición fósiles son organismos extintos que combinan las características de grupos más antiguos y evolutivamente más jóvenes. Permiten identificar vínculos familiares que prueban el desarrollo histórico de la vida. Estas formas se establecen tanto entre animales como entre plantas. La forma de transición de los peces con aletas lobuladas a los anfibios antiguos, los estegocéfalos, es el Ichthyostega. La conexión evolutiva entre reptiles y aves puede ser establecida por el primer pájaro (Archaeopteryx). El vínculo entre reptiles y mamíferos es el lagarto del grupo de los terápsidos. Entre las plantas, la forma de transición de algas a esporas superiores son las psilofitas (las primeras plantas terrestres). El origen de las gimnospermas a partir de pteridofitos lo demuestran los helechos semilleros y el origen de las angiospermas a partir de gimnospermas, de las cícadas.
Serie filogenética (del griego phylon - género, tribu, génesis - origen): secuencias de formas fósiles que reflejan el desarrollo histórico de las especies modernas (filogenia). Actualmente, estas series son conocidas no sólo para los vertebrados, sino también para algunos grupos de animales invertebrados. El paleontólogo ruso V. O. Kovalevsky restauró la serie filogenética del caballo moderno
2. ¿Qué tipos de plantas y animales antiguos conoces?
Respuesta. Hace exactamente 75 años, frente a las costas del sur de África, se descubrió el pez más antiguo del mundo: el celacanto, que existió en la Tierra hace cientos de millones de años. En honor a este evento, te invitamos a aprender sobre ella y otros animales y plantas antiguos que habitan hoy nuestro planeta.
Anteriormente se creía que estos peces se extinguieron a finales del Cretácico (hace 100,5 - 66 millones de años), pero en diciembre de 1938, la curadora del Museo del Este de Londres (Sudáfrica), Marjorie Courtney-Latimer, descubrió un pez con escamas duras y aletas inusuales. en las capturas de los pescadores locales. Más tarde resultó que este pez vivió hace cientos de millones de años y es un fósil viviente.
Desde que este celacanto fue descubierto en el río Chalumna, recibió el nombre de Latimeria chalumnae. Y en septiembre de 1997, en las aguas cercanas a la ciudad de Manado, ubicada en la costa norte de la isla de Sulawesi, los científicos notaron una segunda especie de estos peces: Latimeria menadoensis. Según estudios genéticos, estas especies se separaron hace 30-40 millones de años, pero las diferencias entre ellas son pequeñas.
2. Ginkgo biloba.
En la naturaleza, esta planta crece sólo en el este de China. Sin embargo, hace 200 millones de años se distribuía por todo el planeta, especialmente en el hemisferio norte, en zonas de clima templado y alta humedad. En la Siberia de los períodos Jurásico y Cretácico Inferior había tantas plantas de la clase Ginkgo que sus restos se encuentran en la mayoría de los depósitos de esos períodos. Según los investigadores, en el otoño de aquella época la tierra estaba literalmente cubierta de hojas de ginkgo, como una alfombra.
3. El pequeño ciervo, o kanchil, no sólo es el más pequeño (su altura a la cruz no supera los 25 centímetros y su peso máximo es de unos 2,5 kilogramos), sino también la especie de artiodáctilo más antigua de la Tierra. Estos animales existieron hace 50 millones de años, justo cuando comenzaron a formarse órdenes de ungulados antiguos. Desde entonces, la kanchila se ha mantenido casi sin cambios y se parece más a sus ancestros antiguos que a otras especies.
4. Mariscos de Mississippi.
El marisco del Mississippi, un pez parecido a un caimán, es uno de los peces más antiguos que viven en la Tierra en la actualidad. En la era Mesozoica, sus antepasados habitaron muchos cuerpos de agua. Hoy en día, los mariscos del Mississippi viven en el valle inferior del río Mississippi, así como en algunos lagos de agua dulce de los Estados Unidos.
Estos pequeños crustáceos de agua dulce se consideran las criaturas más antiguas que viven en la Tierra en la actualidad. Los representantes de esta especie apenas han cambiado desde el período Triásico. En aquella época acababan de aparecer los dinosaurios. Hoy en día, estos animales viven en casi todos los continentes excepto en la Antártida. Sin embargo, la especie Triops cancriformis es más común en Eurasia.
6. Metasequoia glyptostroboides.
Estas coníferas estuvieron muy extendidas por todo el hemisferio norte desde el Cretácico hasta el Neógeno. Sin embargo, hoy en día la metasequoia sólo se puede ver en estado salvaje en el centro de China, en las provincias de Hubei y Sichuan.
7. Tiburón duende.
El género Mitsukurina, al que pertenece esta especie de tiburón, se conoció por primera vez a través de fósiles que datan del Eoceno Medio (hace unos 49-37 millones de años). La única especie viva de este género, el tiburón duende, que vive en los océanos Atlántico e Índico, ha conservado algunas características primitivas de sus antiguos parientes y hoy es un fósil viviente.
Preguntas posteriores al § 61
1. ¿Qué es la macroevolución? ¿Qué tienen en común la macroevolución y la microevolución?
Respuesta. La macroevolución es una evolución supraespecífica, a diferencia de la microevolución, que ocurre dentro de una especie, dentro de sus poblaciones. Sin embargo, no existen diferencias fundamentales entre estos procesos, ya que los procesos macroevolutivos se basan en los microevolutivos. En la macroevolución actúan los mismos factores: la lucha por la existencia, la selección natural y la extinción asociada. La macroevolución, como la microevolución, es de naturaleza divergente.
La macroevolución ocurre durante períodos de tiempo históricamente vastos, por lo que es inaccesible a un estudio directo. A pesar de esto, la ciencia tiene mucha evidencia que indica la realidad de los procesos macroevolutivos.
2. ¿Qué evidencia de macroevolución nos aportan los datos paleontológicos? Dé ejemplos de formas de transición.
Respuesta. La paleontología estudia los restos fósiles de organismos extintos y determina sus similitudes y diferencias con los organismos modernos. Los datos paleontológicos nos permiten aprender sobre la flora y la fauna del pasado, reconstruir la apariencia de organismos extintos y descubrir conexiones entre los representantes antiguos y modernos de la flora y la fauna.
Una comparación de restos fósiles de capas terrestres de diferentes eras geológicas proporciona evidencia convincente de los cambios en el mundo orgánico a lo largo del tiempo. Nos permite establecer la secuencia de origen y desarrollo de diferentes grupos de organismos. Por ejemplo, en los estratos más antiguos se encuentran restos de representantes de tipos de animales invertebrados, y en estratos posteriores se encuentran restos de cordados. Incluso estratos geológicos más jóvenes contienen restos de animales y plantas pertenecientes a especies similares a las modernas.
Los datos paleontológicos proporcionan una gran cantidad de material sobre las sucesivas conexiones entre varios grupos sistemáticos. En algunos casos fue posible establecer formas de transición entre grupos de organismos antiguos y modernos, en otros fue posible reconstruir series filogenéticas, es decir, series de especies que se reemplazan sucesivamente entre sí.
En las orillas del norte de Dvina se encontró un grupo de reptiles de dientes salvajes. Combinaron las características de mamíferos y reptiles. Los reptiles con dientes de animales son similares a los mamíferos en la estructura del cráneo, la columna y las extremidades, así como en la división de los dientes en caninos, incisivos y molares.
El descubrimiento de Archaeopteryx es de gran interés desde el punto de vista evolutivo. Este animal del tamaño de una paloma tenía las características de un pájaro, pero también conservaba las características de los reptiles. Signos de aves: extremidades traseras con tarso, presencia de plumas, apariencia general. Signos de reptiles: una larga hilera de vértebras caudales, costillas abdominales y presencia de dientes. Archaeopteryx no podría ser un buen volador, ya que su esternón (sin quilla), los músculos pectorales y los músculos de las alas están poco desarrollados. La columna y las costillas no eran un sistema esquelético rígido y estable durante el vuelo, como en las aves modernas. Archaeopteryx puede considerarse una forma de transición entre reptiles y aves. Las formas de transición combinan simultáneamente las características de grupos antiguos y evolutivamente más jóvenes. Otro ejemplo son los ictiostegas, una forma de transición entre los peces de agua dulce con aletas lobuladas y los anfibios.
3. ¿Cuál es la importancia de reconstruir series filogenéticas?
Respuesta. Serie filogenética. Para varios grupos de animales y plantas, los paleontólogos pudieron recrear una serie continua de formas desde lo antiguo hasta lo moderno, reflejando sus cambios evolutivos. El zoólogo ruso V. O. Kovalevsky (1842-1883) recreó la serie filogenética de los caballos. En los caballos, a medida que pasaban a correr rápido y largo, el número de dedos de sus extremidades disminuía y al mismo tiempo aumentaba el tamaño del animal. Estos cambios fueron consecuencia de cambios en el estilo de vida del caballo, que pasó a alimentarse exclusivamente de vegetación, en busca de la cual era necesario recorrer largas distancias. Se cree que todas estas transformaciones evolutivas tardaron entre 60 y 70 millones de años.
El estudio de series filogenéticas construidas sobre la base de datos de paleontología, anatomía comparada y embriología es importante para el desarrollo posterior de la teoría general de la evolución, la construcción de un sistema natural de organismos y la reconstrucción de la imagen de la evolución de un grupo sistemático específico de organismos. Actualmente, para construir series filogenéticas, los científicos utilizan cada vez más datos de ciencias como la genética, la bioquímica, la biología molecular, la biogeografía, la etología, etc.
Uno de los más conocidos y estudiados es la serie filogenética de los ungulados modernos de un solo dedo. Múltiples hallazgos paleontológicos y formas de transición identificadas crean una base de evidencia científica para esta serie. La serie filogenética del caballo, descrita por el biólogo ruso Vladimir Onufrievich Kovalevsky en 1873, sigue siendo hoy un “icono” de la paleontología evolutiva.
Evolución a través de los siglos
En la evolución, las series filogenéticas son formas de transición sucesivas que llevaron a la formación de especies modernas. Según el número de enlaces, la serie puede ser completa o parcial, pero la presencia de formas de transición sucesivas es un requisito previo para su descripción.
La serie filogenética del caballo se considera evidencia de evolución precisamente por la presencia de formas secuenciales que se reemplazan entre sí. La multiplicidad de hallazgos paleontológicos le confiere un alto grado de fiabilidad.
Ejemplos de series filogenéticas.
La fila de caballos no es la única entre los ejemplos descritos. La serie filogenética de ballenas y aves ha sido bien estudiada y tiene un alto grado de confiabilidad. Y controvertida en los círculos científicos y más utilizada en diversas insinuaciones populistas es la serie filogenética de los chimpancés y los humanos modernos. En la comunidad científica continúan las disputas sobre los eslabones intermedios que faltan aquí. Pero no importa cuántos puntos de vista haya, la importancia de las series filogenéticas como evidencia de la adaptabilidad evolutiva de los organismos a las condiciones ambientales cambiantes sigue siendo indiscutible.

La conexión entre la evolución de los caballos y el medio ambiente.
Múltiples estudios realizados por paleontólogos han confirmado la teoría de O. V. Kovalevsky sobre la estrecha relación de los cambios en el esqueleto de los antepasados de los caballos con los cambios en el medio ambiente. El clima cambiante provocó una disminución de las superficies forestales y los antepasados de los ungulados modernos de un solo dedo se adaptaron a las condiciones de vida en las estepas. La necesidad de movimientos rápidos provocó modificaciones en la estructura y número de dedos de las extremidades, cambios en el esqueleto y los dientes.
El primer eslabón de la cadena.
A principios del Eoceno, hace más de 65 millones de años, vivió el primer antepasado del caballo moderno. Se trata de un “caballo bajo” o Eohippus, que era del tamaño de un perro (hasta 30 cm), descansaba sobre todo el pie de una extremidad, que tenía cuatro (delanteros) y tres (posteriores) dedos con pequeñas pezuñas. Eohippus se alimentaba de brotes y hojas y tenía dientes tuberculados. Color marrón y pelo escaso en una cola móvil: este es el ancestro lejano de los caballos y las cebras en la Tierra.
Intermedios
Hace unos 25 millones de años, el clima del planeta cambió y las extensiones de estepa comenzaron a reemplazar a los bosques. En el Mioceno (hace 20 millones de años) aparecieron mesohippus y parahippus, más parecidos a los caballos modernos. Y se considera que el primer ancestro herbívoro de la serie filogenética del caballo es Merikhippus y Pliohippus, que entraron en la arena de la vida hace 2 millones de años. Hipparion: el último eslabón de tres dedos
Este ancestro vivió en el Mioceno y Plioceno en las llanuras de América del Norte, Asia y África. Este caballo de tres dedos, parecido a una gacela, aún no tenía pezuñas, pero podía correr rápido, comía hierba y era ella quien ocupaba vastos territorios.

Caballo de un dedo - pliohyppus
Estos representantes de un dedo aparecieron hace 5 millones de años en los mismos territorios que los hippariones. Las condiciones ambientales están cambiando: se están volviendo aún más secas y las estepas se están expandiendo significativamente. Aquí es donde el uso de un solo dedo resultó ser un signo más importante para la supervivencia. Estos caballos medían hasta 1,2 metros de altura a la cruz, tenían 19 pares de costillas y fuertes músculos en las piernas. Sus dientes adquieren largas coronas y pliegues de esmalte con una capa de cemento desarrollada.
caballo que conocemos
El caballo moderno, como etapa final de la serie filogenética, apareció al final del Neógeno, y al final de la última glaciación (hace unos 10 mil años), millones de caballos salvajes ya pastaban en Europa y Asia. Aunque los esfuerzos de los cazadores primitivos y la reducción de los pastos hicieron que el caballo salvaje fuera una rareza hace ya 4 mil años. Pero dos de sus subespecies, el tarpán en Rusia y el caballo de Przewalski en Mongolia, lograron sobrevivir mucho más que todos los demás.

Caballos salvajes
Hoy en día prácticamente no quedan verdaderos caballos salvajes. El tarpan ruso se considera una especie extinta y el caballo de Przewalski no se encuentra en condiciones naturales. Las manadas de caballos que pastan libremente son formas salvajes domesticadas. Aunque estos caballos regresan rápidamente a la vida salvaje, todavía se diferencian de los caballos verdaderamente salvajes.
Tienen melenas y colas largas y son de diferentes colores. Exclusivamente los caballos pardos de Przewalski y sus tarpans ratoncitos tienen flequillos, crines y colas recortados.

En América Central y del Norte, los caballos salvajes fueron completamente exterminados por los indios y aparecieron allí sólo después de la llegada de los europeos en el siglo XV. Los salvajes descendientes de los caballos de los conquistadores dieron lugar a numerosas manadas de mustangs, cuyo número hoy se controla mediante disparos.
Además de los mustangs, en América del Norte hay dos especies de ponis insulares salvajes: en las islas Assateague y Sable. En el sur de Francia se encuentran manadas semisalvajes de caballos de Camarga. También se pueden encontrar algunos ponis salvajes en las montañas y páramos de Gran Bretaña.

Nuestros caballos favoritos
El hombre domó al caballo y crió más de 300 de sus razas. Desde pesos pesados hasta ponis en miniatura y hermosos caballos de carreras. En Rusia se crían unas 50 razas de caballos. El más famoso de ellos es la manita de Oryol. Bata exclusivamente blanca, excelente trote y agilidad: estas cualidades fueron tan valoradas por el Conde Orlov, considerado el fundador de esta raza.